

Introduction
전세계적으로 인간의 활동에 의한 기후변화 및 도시화 등 식물의 환경 요인이 급변하고 있다. 특히, 도심지에서 생장하는 식물은 인공적인 생육 환경 (온도, 수분, 토양 및 광 환경 등)에서 재배되고 있어 식물의 발아, 개화, 엽면적, 기공 개폐, 엽록소 함량 및 광합성 능력 등에 영향을 미친다 (Cho et al., 2008; Kang et al., 2016). 이러한 환경 요인은 생육 시기별 및 계절적 변화에 따라 달라지므로 비파괴적 방법을 통한 지속적인 모니터링이 필요하다. 본 연구는 비파괴적인 엽록소 형광 (OJIP) 분석을 이용한 도심지 토양에 식재된 지피식물의 건전성 평가 및 내성식물 선발 연구를 목적으로 하였다.
지금까지 쾌적한 도시환경과 스마트시티 건설 사업으로 도로와 주차장 등의 시설들이 지하로 배치면서, 대규모의 인공지반이 조성되고 생태적 환경 조성을 위한 녹지면적은 증가하고 있지만. 인공지반에 조성된 토양 대부분은 여러 종류의 토양이 섞인 상태에서, 중장비 등 답압의 영향으로 불투수층이 됨으로서 배수가 불량한 상태가 되고 생육부진, 고사 등으로 도시환경과 미관을 오히려 해치는 문제점들이 발생하고 있다 (Kays, 2013). 이에 대한 대책으로 암거배수, 객토, 유효토층의 개량 등의 토양 관리 계획과 함께 표토층을 피복하면서 뿌리를 내려 토양의 물리성을 개량하고 토양을 비옥하게 만드는 지피식물의 생태적 가치가 중요하게 부각되고 있다 (Park, 2017). 지피식물의 식재 등 생태적인 복원에 관심이 집중되고 있다.
그러나 현재 지피식물의 식재현황을 조사한 바에 따르면 맥문동, 옥잠화, 비비추, 잔디를 제외하고는 식재빈도가 매우 적어 지피식물이 다양하게 식재되지 못하고 잔디 단일 품종으로 식재하거나, 3 - 4 품종이 다수의 장소에 식재되고 있는데 (Kim, 2004), 잔디피복은 사후관리 비용이 많이 발생하고, 광요구도가 높은 특성으로 건물 음영에 의한 음지에서는 고사하여 나지를 형성하게 되는 문제가 빈번히 발생하고 있다.
일반적으로 식물은 환경 변화에 의해 광합성 과정 중 전자전달 저해, 엽록소 함량 변화, 전하분리 감소 등 광합성 효율이 감소하며 (Oukarroum et al., 2007; Perboni et al., 2012), 식물체 내 단백질 변화에 의한 빛의 반사지수가 다르게 나타나는 것으로 밝혀졌다 (Rouse et al., 1974; Gamon et al., 1992; Suárez et al., 2009).
최근 비파괴적으로 모니터링이 가능한 엽록소 형광 분석법을 통해 환경 스트레스에 의한 식물의 건전성 평가에 엽록소 형광 분석법이 염해, 한발, 고온 및 저온 등 다양한 환경 조건에서 활용되고 있다 (Strasser, 1985; Govindjee, 1995; Prakash et al., 2003; Yoo et al., 2014). 광환경 변화와 관련하여 피음도가 증가함에 따라 엽록소 a/b ratio가 감소한 것으로 보고되었으며 (Cho et al., 2008), 내음성 식물 선발에 엽록소 형광 분석법을 활용한 광화학 성능지수 (PI) 등 광생리 지표 연구가 수행되었다 (Kang et al., 2016). 본 연구는 차광 조건에서 식물의 내음성 특성에 따라 엽록소 a/b ratio 및 광합성 색소 함량 변화가 발생해 결국 광 이용 효율의 차이가 나타날 것으로 예측되어 엽록소 형광 분석을 통해 광 부족에 의한 스트레스 평가를 실시하고자 하였다.
그리고 일조부족, 건조 등 도시환경 특성을 고려하여 내음성 지피식물 식재설계 기술로 일조환경 차이에 의한 식물의 생리ㆍ생장 특성을 파악하여 불규칙적인 도시 광환경 특성에 맞는 식물을 선발하는 방법으로 광화학 해석 연구를 수행하였다.
Materials and Methods
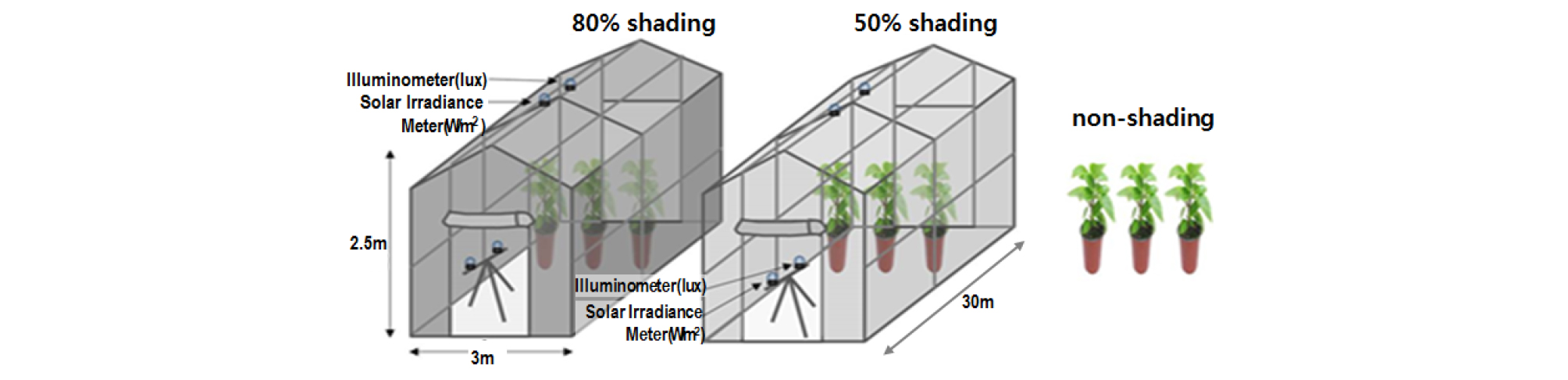
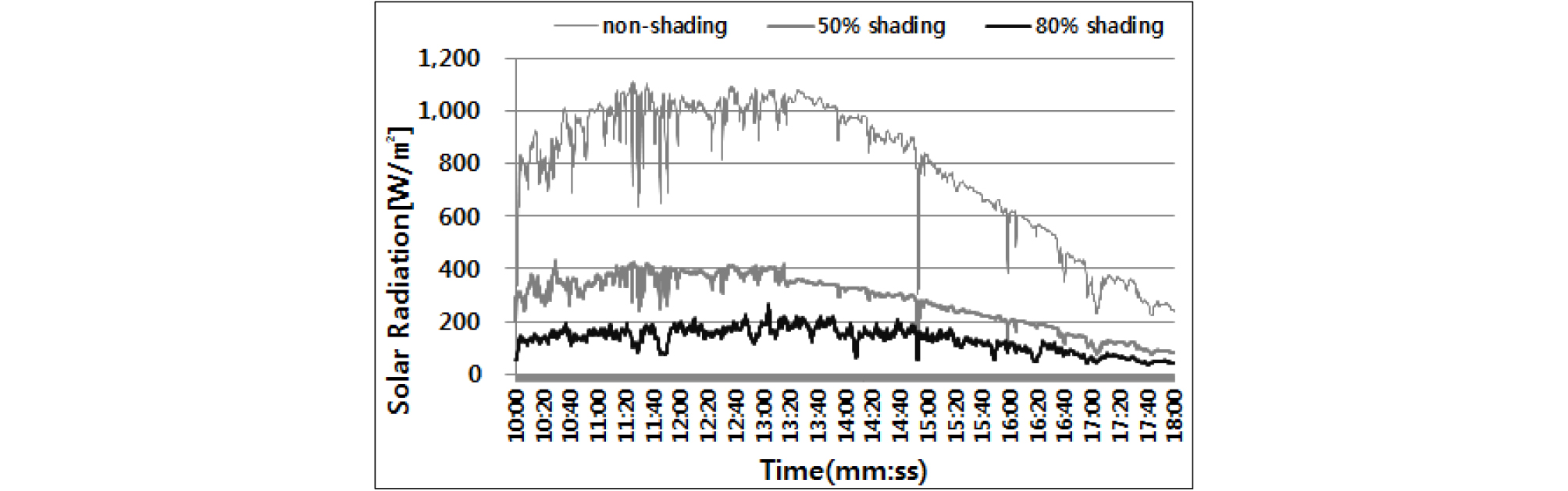
아파트 조경공간 나지에 의한 식재불량지역의 일조 특성을 분석한 결과 건물에 의한 영구음지 지역은 제외하고 식재 가능한 지역 중 가장 낮은 식재공간은 하루 2시간 이상 간헐적 일조가 유입되는 지역으로, 지속광 조건으로 환산하면 노지의 15 - 20% 일조유입 유형으로 구분하였다. 또한 건물과의 이격거리가 5m - 10m의 낙엽활엽교목 하부는 노지대비 50 - 20%의 일조차폐율을 나타냈다 (RDA, 2017). 선행연구 결과를 기준으로 지피식재를 위한 차광시험은 자연광의 50%, 80% 일조차폐율 환경 조건에서 실시하였는데 노지 공간과 차광률이 50%, 80%인 차광막을 온실 지붕에 설치하고 (Fig. 1) 조도센서 (AHLBORN, FLA646-E1)를 사용하여 시험구내 조도 분포를 모니터링하였다 (Fig. 2).
침식과 건조로부터 표토를 보호하고 잡초의 침입을 억제하여 나지를 피복하여 경관의 심미성을 향상시키기 위한 지피식물로는 광엽성 잎으로 지표면을 덮어주거나 빠르게 피복하면서 관상가치를 주는 다년생 초화류가 적합한데 본 실험의 공시식물 6종의 기본 특성과 형상은 Table 1과 같다.
Table 1. Growth and ornamental characteristics of experiment plants.
식재토양은 도시녹지 토양의 특성은 답압으로 토양 경도가 높고 pH가 7.2 - 7.4로 약알칼리성으로 유기물함량이 1.0 - 1.5%의 매우 낮은 환경으로 (Kil, 2001), 묘종의 뿌리활착, 배수 및 통기성 유지를 위한 식재지반을 개선하기 위하여 인공상토 (버미큘라이트:펄라이트:피트모스:라임스톤 = 1:1:1:1, Sun Gro Horticulture, Sunshine® Mix #4)를 사용하였다.
실험식물의 차광처리에 의한 생장 특성의 시계열 변화를 분석하기 위하여 15일 간격으로 식물의 초장, 마디수, 옆폭, 엽장을 측정하고, SPSS ver.20을 이용하여 차광처리별 생장 차이는 Duncan의 다중검정 (multiple rage test)를 사용해서 5% 유의수준으로 검증하였고 차광 기간 경과에 따른 생장 요인의 변화는 단계별선택법 (stepwise selection method)을 활용하여 측정항목 간의 다중공선성을 검토하였다.
식물의 차광 조건에 따른 일조 스트레스는 엽록소 형광 반응 (OJIP)을 활용해 광생리지표를 분석하였는데, 식물체 잎을 leaf clip로 이용하여 30분간 암처리를 한 후 Fluorpen FP-100 (PSI, Czech)으로 OJIP를 3반복 측정하였다. 측정된 엽록소 형광매개변수는 Stirbet and Govindjee (2011)의 방법에 의하여 JIP-Test를 실시하고 (Table 2) ANOVA 분석을 통계적 유의성을 검증하였다.
Table 2. Equations and definitions of chlorophyll fluorescence parameters (modified from Stribet and Govindjee, 2011).
Results and Discussion
일반적으로 식물은 생육상태가 좋을 때 초장, 엽면적 등이 성장하지만 차광정도가 높아짐에 따라 초장, 엽장 등의 길이는 길어진다. 하지만 건강한 생장조건에서는 엽의 밀도가 높아지지만 차광조건에서는 엽수, 분얼수 등 엽의 밀도도 함께 줄어드는 변화를 보이는데, 이는 광 부족에 의해 생육 발달이 억제되고 엽록소의 변화로 엽색이 짙어지며, 영양생장에서 생식생장 전환이 늦어지는 등의 생육변화에 의해 유발되는 외부적인 특성이다 (Kim et al., 2001).
따라서 스트레스에 대한 다양한 형태적인 변화를 유발하는 식물 생리적인 특성을 조사함으로 스트레스 상태인지 생장하는 상태인지를 정량적으로 판단하고, 보다 정확한 내음성 특징을 조사하기 위하여 본 연구에서는 엽록소 형광반응 (OJIP)분석을 통해 차광처리에 따른 정원 식물의 광 이용효율을 평가 및 분석하였다.
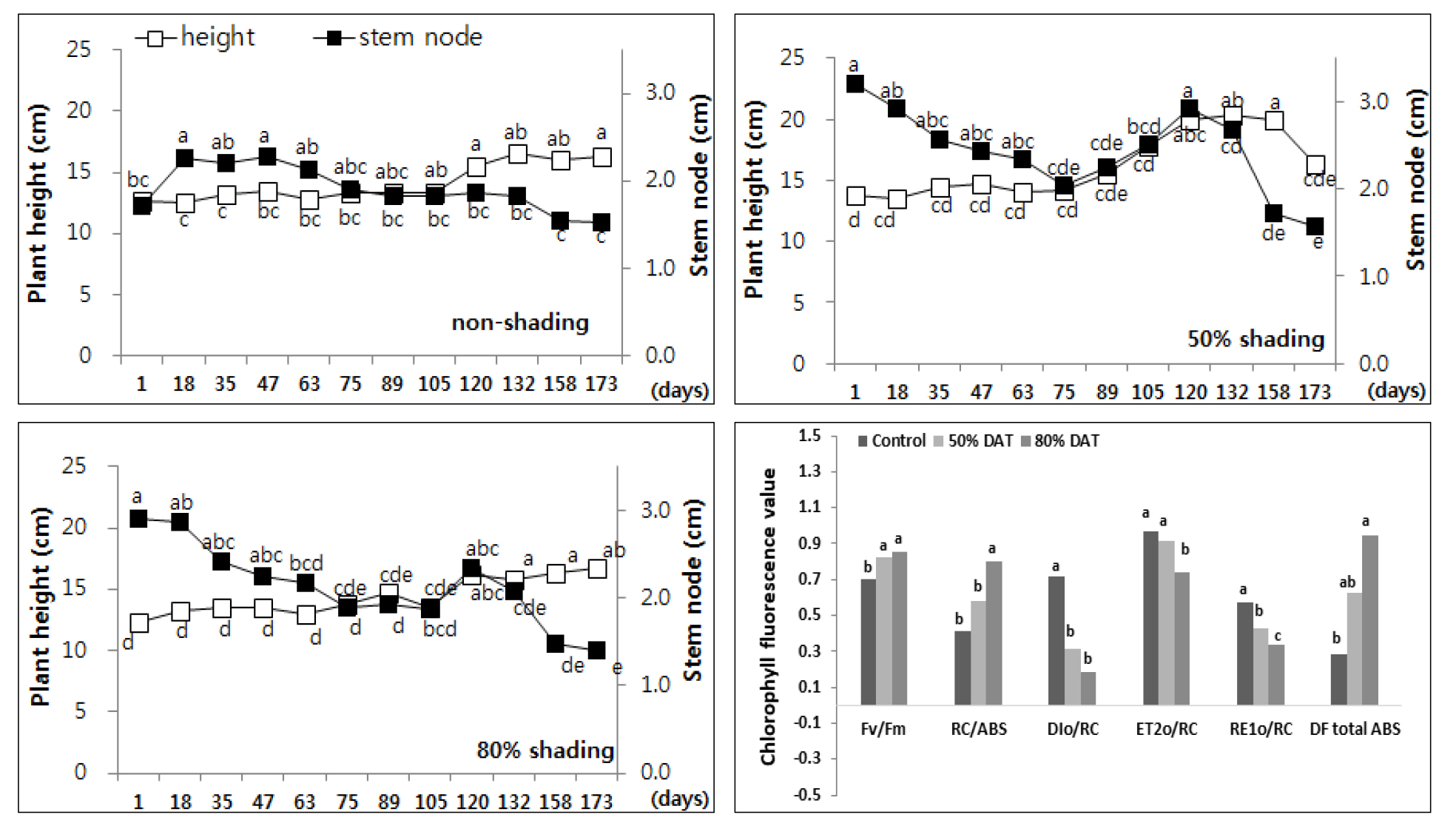
일반적으로 자생지 환경이 양지/반음지 조건에서 50 cm까지 자라는 꼬리풀은 (Lee, 2004) 차광처리를 하지 않은 곳에서는 초장이 37.9 ± 0.89 cm, 50% 차광구에서는 61.67 ± 6.83 cm, 80% 차광에 의해 75.5 ± 1.95 cm 생장하였다(Fig. 3).
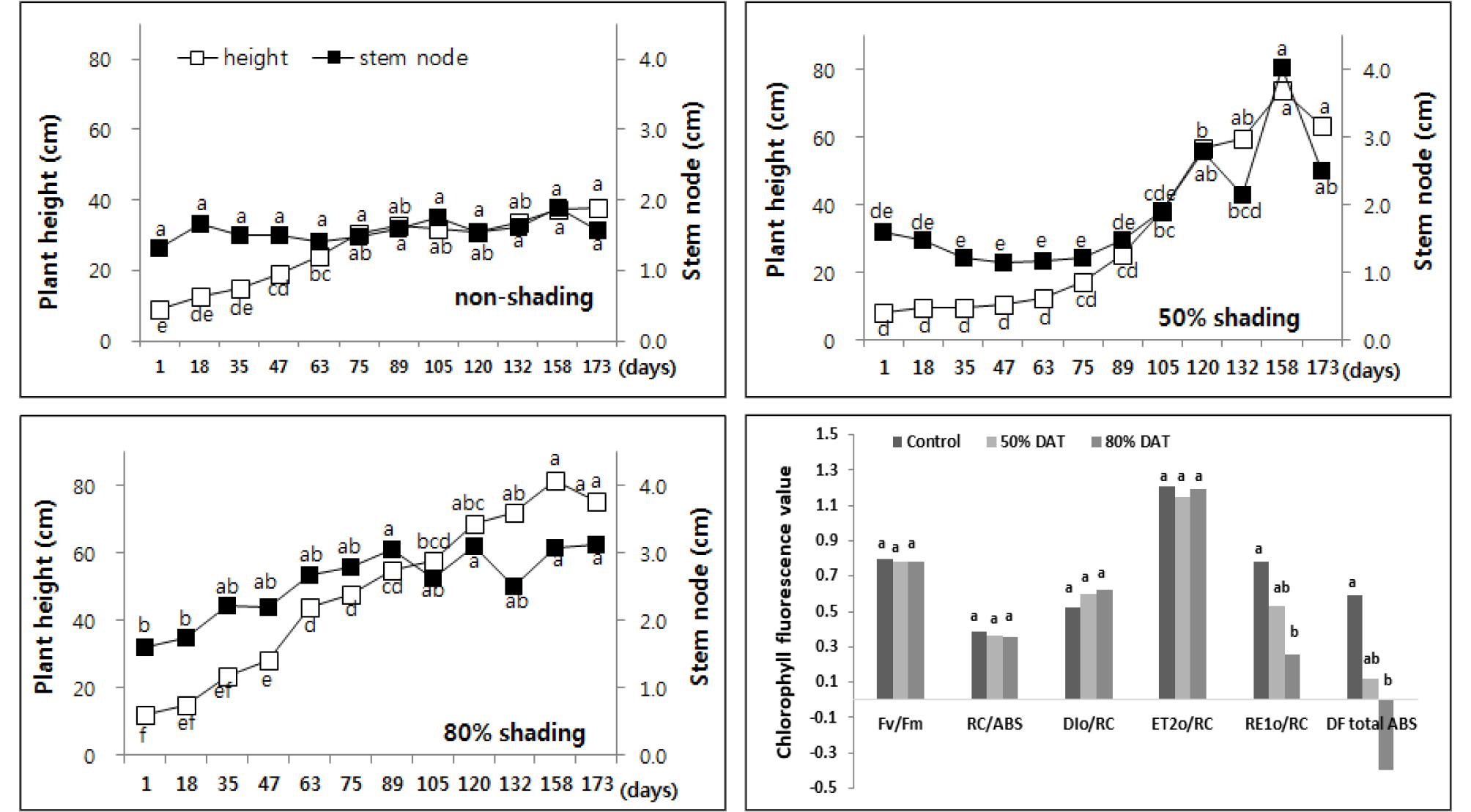
Fig. 3.
Changes of plant height and length of stem node every two weeks of Veronica linariifolium by non shading (top left), 50% shading (top right), and 80% shading (bottom left) treatment relatively. Different letters in one measurement indicate statistically significant difference at p ≤ 0.05 by Duncan multiple range test.
꼬리풀 (Veronica linariaefolia)은 초장은 계속 증가하면서도 마디 길이 변화는 없는 것으로 보아 차광에 의한 생장이 증가하여 내음성 식물로 가치가 있는 종으로 나타났다.
엽록소 형광 분석 결과 꼬리풀 등 6종의 모든 식물에서 광합성계 II 최대 양자수득률 (FV/FM Ratio)의 변화가 없거나 차광구에서 높은 것으로 확인되었다. 꼬리풀은 차광 조건에서 광합성계 II 전자전달 에너지 플럭스 (ET2o/RC)는 변화가 없었으나 광합성계 I 전자전달 에너지 플럭스 (RE1o/RC)와 식물의 활력을 나타내는 DFtotal ABS 가 차광률이 증가할수록 크게 감소하였다 (Fig. 3).
일반적으로 RC/ABS는 빛에 의한 광화학 기구의 개폐와 관련하여 광합성계 II 전자전달 과정 중 QA의 환원 (QA → QA-) 정도를 확인할 수 있는 형광 매개변수로 알려져 있다. 본 연구에서 꼬리풀은 노지에 비해 50% 와 80% 차광구에서 RC/ABS 가 각각 6% 와 8% 감소함에 따라 광합성계 II 반응중심당 에너지 소실 (DIo/RC)이 15% - 20% 증가하는 것으로 확인되었다.
꼬리풀과 같이 수직으로 생장하는 꽃범의꼬리 (Physostegia virginiana)도 50%, 80%로 차광률이 높아질수록 초장은 51%, 44% 증가하였으나 마디 길이는 감소였다. 이와 같은 결과는 상대적인 성장 지표를 나타내는 마디수의 중가를 의미하므로 음지에서 더 안정적인 생장을 나타내는 종으로 꼬리풀에 비해 더 높은 차광환경에서도 식재 가능할 것으로 사료되었다 (Fig. 4).
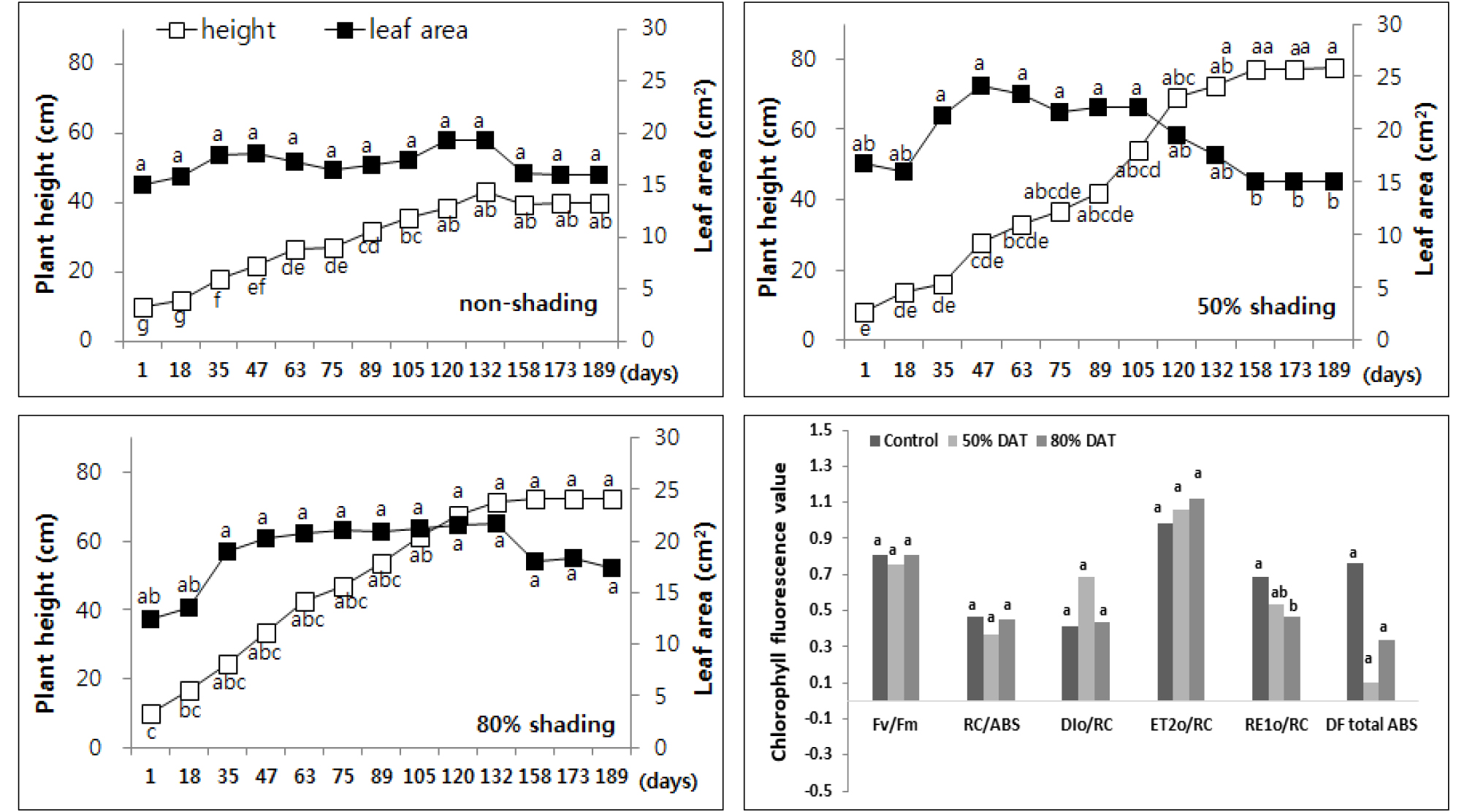
Fig. 4.
Changes of plant height and leaf area every two weeks of Physostegia virginiana by non shading (top left), 50% shading (top right), and 80% shading (bottom left) treatment relatively. Different letters in one measurement indicate statistically significant difference at p ≤ 0.05 by Duncan multiple range test.
그리고 엽록소 형광 분석 결과에서 꼬리풀과 유사한 경향을 보이며 나지에 비해 차광 조건에서의 광 이용 효율이 낮았으나 차광률이 높아질수록 생장량이 증가했던 결과와 다르게 50% 차광구에서 RE1o/RC를 제외한 5종의 형광 매개변수의 광 이용 효율이 80% 차광구 보다 낮은 것으로 확인되었다. 특히 50% 차광구에서 RC/ABS가 약 21% 감소하며 활성화된 광합성계 II 반응중심당 양자 흡수량이 크게 증가해 손실되는 양자 비율 (DIo/RC)이 상대적으로 크게 증가하며 광 이용 효율이 저하된 것으로 사료되었다.
반구형으로 자라는 무늬사초 (Carex maculata)도 노지에 비해 50% 차광구에서 엽폭, 엽장 길이의 차이 없이 초장 신장이 가장 좋았고, 엽록소 형광 분석 결과에서도 무늬사초가 6종의 식물 중 가장 높은 광 이용 효율을 보이며 노지에 비해 차광환경이 생장에 좋은 조건인 것으로 사료되었다 (Fig. 5). 차광 조건에서 광합성계 II 의 활성 (RC/ABS)이 17% 이상 증가하며 열에 의한 에너지 손실 (DIo/RC)이 노지에 비해 30% 가까이 감소하는 결과를 보이며 광합성계 II에서 광합성계 I 로의 전자전달 에너지 플럭스가 높게 유지되는 것으로 판단되었다. 따라서 무늬사초는 차광 조건에서 광화학 전자전달 효율이 높아 광합성 명반응의 최종 산물인 NADP 환원 (NADP→NADPH) 및 ATP 합성 (ADP→ATP)이 증가할 것으로 사료되었다.
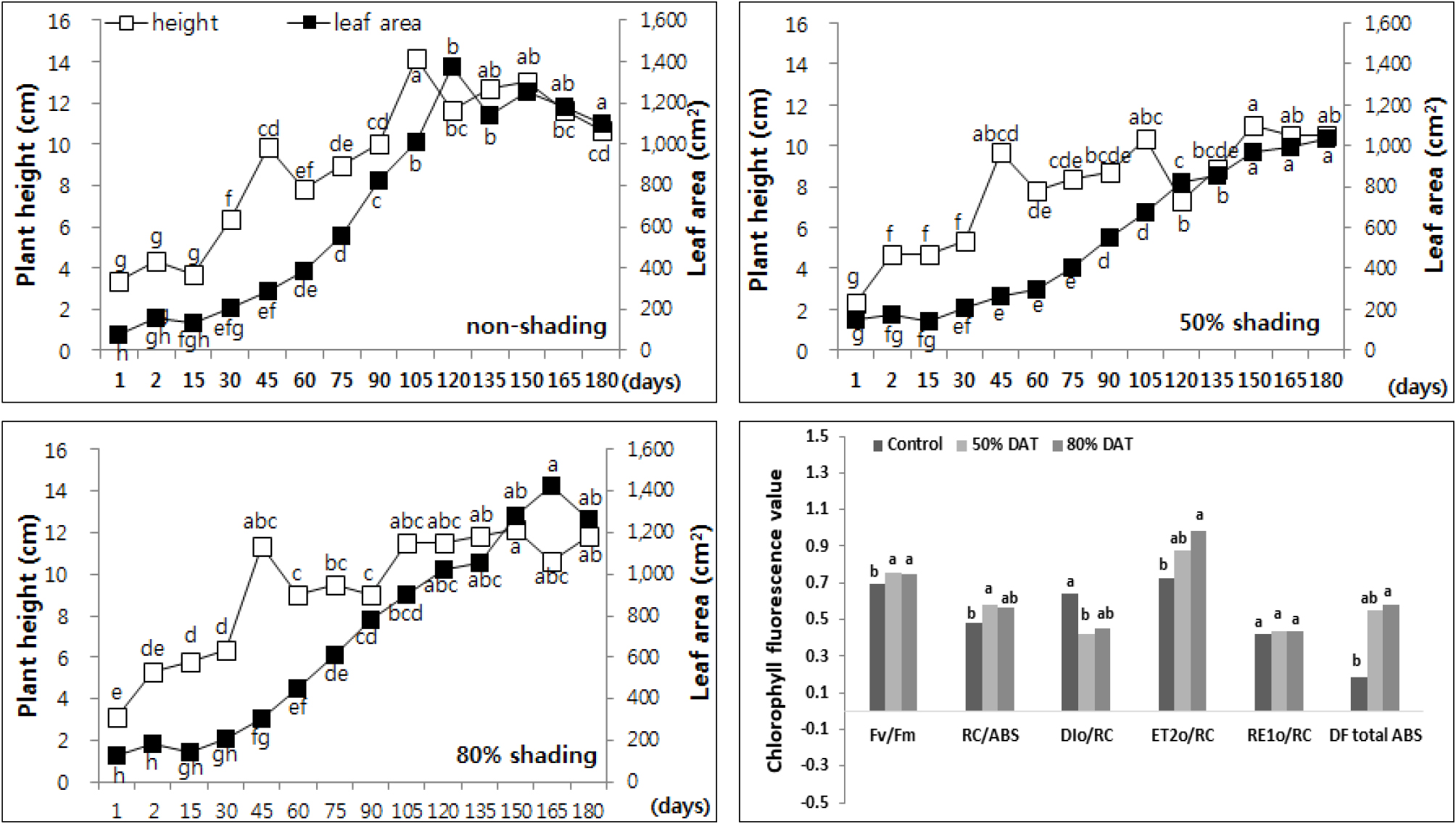
Fig. 5.
Changes of plant height and leaf area every two weeks of Carex maculata by non shading (top left), 50% shading (top right), and 80% shading (bottom left) treatment relatively. Different letters in one measurement indicate statistically significant difference at p ≤ 0.05 by Duncan multiple range test.
회양목과에 속하는 수호초 (Pachysandra terminalis 'Variegata')는 7개월간 시간이 경과함에 따른 성장량이 적어서 차광에 의한 형태적인 변화는 미미했지만 노지에서 4.0 cm 생장한데 비해 50% 차광에서는 6.9 cm로 초장과 마디수가 증가하여 마디길이는 짧아지면서 단단하게 자라 엽의 밀도도 높아져 지표면 피복효과가 좋았다 (Fig. 6). 수호초는 80% 차광구에서 광합성계 II 와 광합성계 I 의 전자전달 효율은 감소했으나 광화학 기구의 활성 측면에서는 노지에 비해 50% 및 80% 차광구에서 광합성계 II 반응중심의 활성도 (RC/ABS)가 각각 39%, 93% 증가했으며, 광화학적 활력 (DFtotal ABS)이 두배 이상 높은 것으로 나타났다. 따라서 차광구 수호초의 광화학 전자전달 효율은 노지에 비해 낮았으나 식물의 활력이 높아 차광 스트레스에 의한 광화학 기구의 저해 현상은 아닌 것으로 사료되었다.
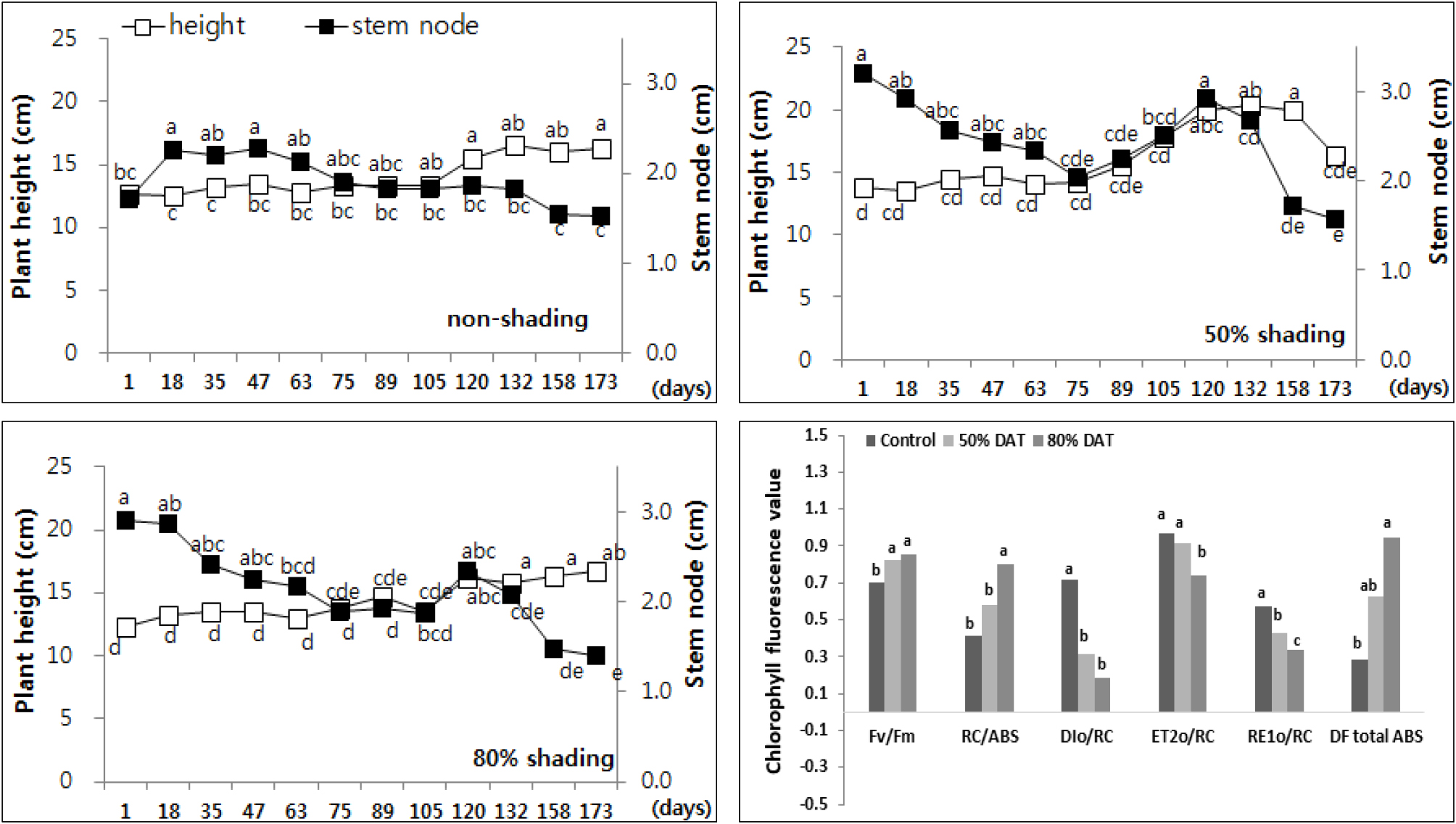
Fig. 6.
Changes of plant height and length of stem node every two weeks of Pachysandra terminalis ‘Variegata’ by non shading (top left), 50% shading (top right), and 80% shading (bottom left) treatment relatively. Different letters in one measurement indicate statistically significant difference at p ≤ 0.05 by Duncan multiple range test.
감국 (Chrysanthemum indicum)의 생장형태는 대부분 덤불형으로 생육초기에는 직립형으로 자라다가 화뢰형성기를 지나면서 마다길이가 급격히 증가하며 초폭이 넓어지는 반직립 또는 반구형으로 자라는데, 정식시기가 빠를수록 착화수와 초장, 화경장이 많고 길어진다 (Jung, 2011). 국화과 식물은 단일조건에서 개화가 촉진되고 장일에서는 영양생장이 촉진되는데, 국화의 광스트레스에 대한 최초의 징후는 광스트레스가 증가함에 따라 영양생장이 지속되어 개화가 지연된다 (De Jong, 1986).
본 연구에서는 50% 차광에서는 차이를 보이지 않다가 80% 차광구에서 초장이 급격하게 증가 (57.4%)하였으며, 엽면적의 변화도 같은 양상을 보였는데 이러한 변화가 생장 요인으로 작용했는지, 스트레스 요인에 의한 변화인지를 확인해본 결과 80% 차광구에서 광 이용 효율의 급격한 감소가 확인되어 차광에 의해 스트레스를 받고 있음을 알 수 있었다 (Fig. 7).
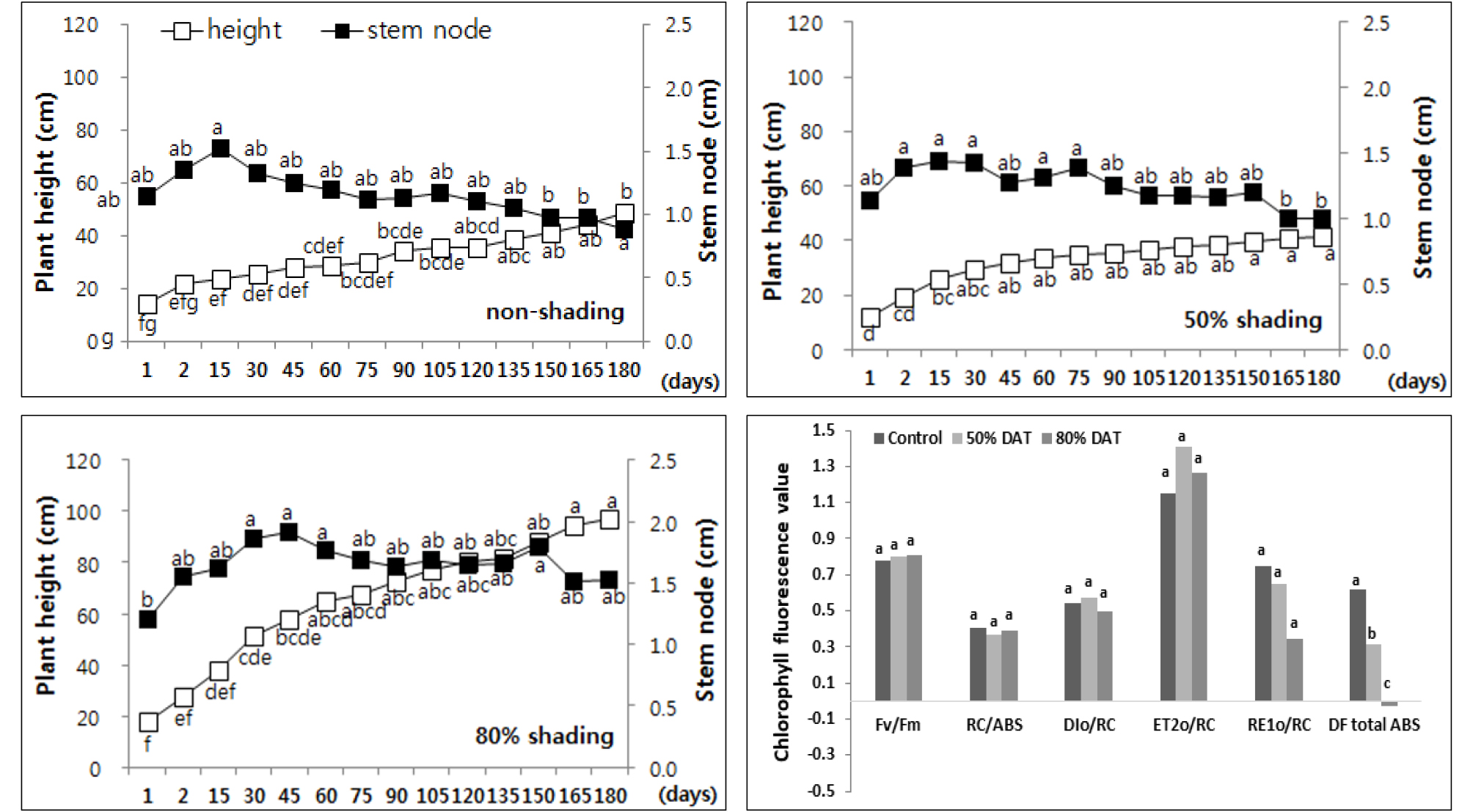
Fig. 7.
Changes of plant height and length of stem node every two weeks of Chrysanthemum indicum by non shading (top left), 50% shading (top right), and 80% shading (bottom left) treatment relatively. Different letters in one measurement indicate statistically significant difference at p ≤ 0.05 by Duncan multiple range test.
감국은 차광 처리에 의해 광계II 암적응 최대양자수득률 (FV/FM)은 변화가 없었으나 FV(FM-FO)가 46% - 60% 증가하며 차광 조건에서 엽록소 함량 증가에 의해 최대 형광량이 크게 증가한 것으로 확인되었다.
감국은 차광 조건에서 광합성계 II 전자전달 에너지 플럭스 (ET2o/RC)는 노지에 비해 높았으나 광합성계 I 전자전달 에너지 플럭스 (RE1o/RC) 및 광화학적 활력 (DFtotal ABS)은 50% 차광구에서 각각 13%, 49% 감소했으며, 80% 차광구에서는 54%, 104% 이상 감소하며 광합성계 I의 비활성화로 인한 전자전달 저해가 나타난 것으로 확인되었다.
따라서 감국은 50% 차광구에서 형태적인 변화가 없었으나 80% 차광구에서 초장 증가 및 광 이용효율 감소는 차광량의 증가에 따른 광스트레스가 나타난 것으로 사료되었다.
노루오줌 (Astilbe rubra)은 봄에 발아해서 초장이 신장하고 6월 - 8월에 개화한 후 2 - 3개월 동안 주 (株)가 비대되고, 11월부터 다음 해 눈의 형성하고 잎이 쇠퇴하면서 휴면에 들어가는 생육상을 가지고 있다 (RDA, 2013). 내한·내서성이 강하고 반그늘의 약간 습기가 있는 곳에서 60 - 80 cm까지 자라는데 직사광선이 닿는 곳에서도 연 1회 개화하는 1계성의 작물로 화아분화에는 겨울에 충분한 저온이 필요하고 일장은 개화에 영향을 끼치지 않는 것으로 보고된다.
4월부터 7개월간의 차광에 의한 생장 차이를 조사한 결과 노지에서는 7월까지 60.4% 초장이 신장하였는데 50% 차광구에서는 초장이 17.2%, 80%에서는 16.7%로 감소하였고, 엽면적이 노지생장에 비해 23.5% 늘어나 차광에 영향을 받고 있는 결과를 보였고 80% 차광에서는 엽면적도 작아져 가시적으로도 생육이 불량한 상태임을 알 수 있었다. 엽록소 형광 분석 결과는 감국과 유사한 경향을 보이며 차광에 의한 광화학 기구의 광 이용 효율이 감소하였다 (Fig. 8). 특히 노루오줌은 50% 와 80% 차광구 모두 광합성계 II 반응중심의 활성 (RC/ABS)은 높게 유지되었으나, 광합성계 II 와 광합성계 I 전자전달 에너지 플럭스 (ET2o/RC, RE1o/RC)가 감소하며 모든 광화학 기구에서 전자전달 저해가 발생하는 것으로 사료되었다.
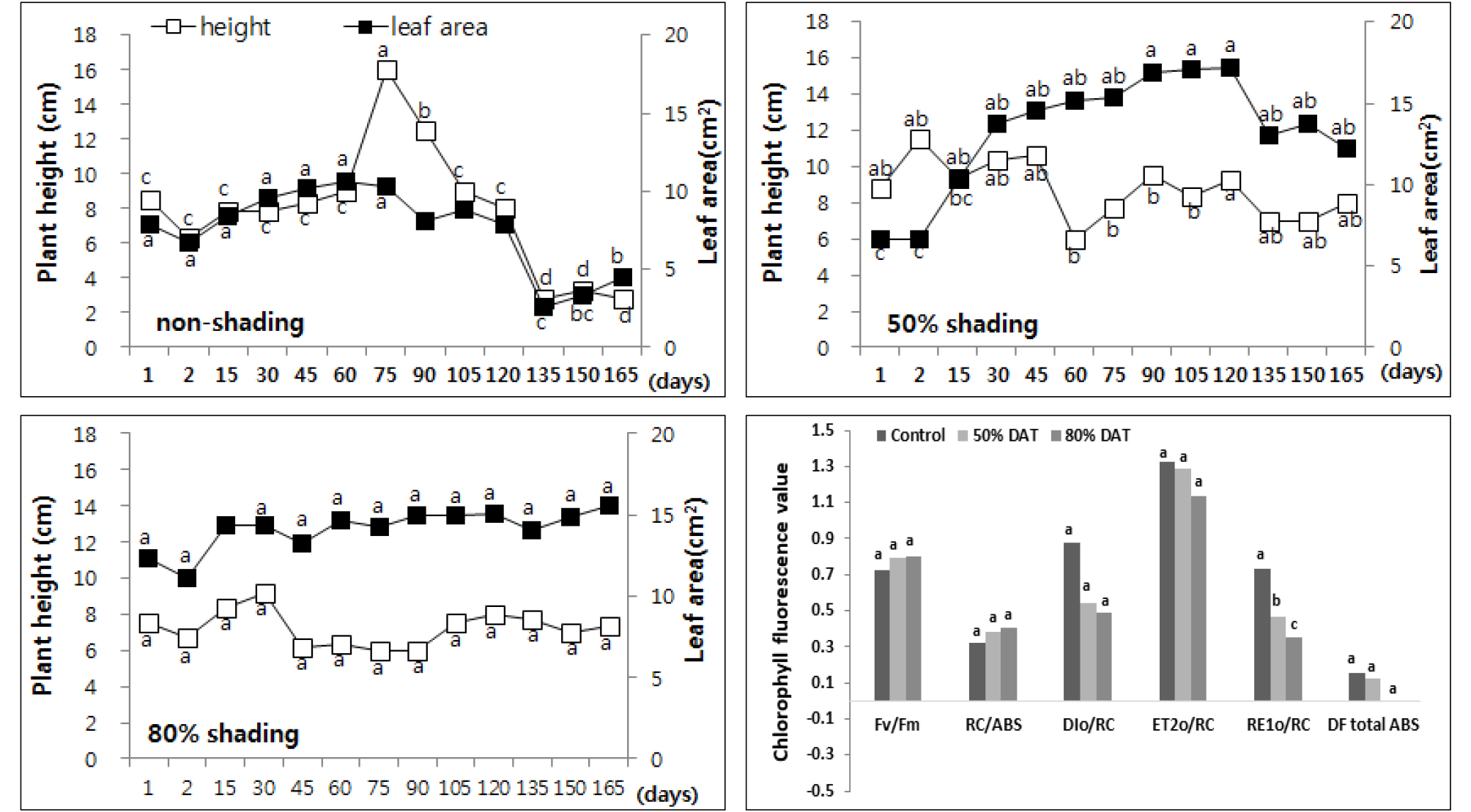
Fig. 8.
Changes of plant height and leaf area every two weeks of Astilbe rubra by non shading (top left), 50% shading (top right), and 80% shading (bottom left) treatment relatively. Different letters in one measurement indicate statistically significant difference at p ≤ 0.05 by Duncan multiple range test.
본 연구에서 지피식재용 경관 식물로 분류되어 있는 국내 자생 초본류 6종을 대상으로 차광에 차이에 의한 형태적인 생장변화를 분석한 결과 모든 식물이 노지에 비해 50% 차광에서 초장이 신장하였는데 이 중 꼬리풀과 꽃범의 꼬리, 무늬사초, 수호초는 마디수가 많아져 차광에 의해 생장이 증가한 반면 감국은 초장과 엽면적은 신장하는데 마디 수는 감소하여 광부족에 의한 웃자람 현상을 보였다. 노루오줌은 초장은 감소하고 엽면적은 늘어나며 광부족 현상에 의한 이상 성장의 형태를 나타냈다.
지피식물 중 가장 많이 식재되고 있는 맥문동을 대상으로 내음성 연구가 많이 진행되었는데 50%차광에서 엽수, 엽면적지수, 현존량, 엽록소 함량 등이 높아지고 (Shin, 1987), 초장과 엽장은 0 - 50%차광까지는 증가하고 그 이상에서는 감소하며, 엽폭은 차광이 높을수록 감소하는 등 (Hyun, 1996) 차광에 의한 형태적인 변화로 본 연구 결과를 유추할 수 있었다. Ding et al. (2005)은 일곱 수준의 빛 강도 (1.6, 3.1, 6.3, 12.5, 25, 50, 100%) 에 따른 왜성 소엽 맥문동의 생장 실험을 통해 25% - 50% 일조량 범위에서 엽록소 함량이 높으며 분얼이 빠르고 뿌리 생명력이 강해지는 것을 확인하였고, Kim (2010)은 왜성 소엽맥문동은 음지에서 엽록소 값이 두배 이상 높아 양지보다 음지에 식재해야 동절기 때 엽색의 탈색을 방지할 수 있다고 보고하였다. Kim (1999)과 Choi et al. (2001)은 차광에 따른 맥문동 무늬종의 잎색 변화를 조사하였는데 노랑무늬 맥문동의 반엽 발현은 차광에 의해 저하됨을 밝히는 등 차광에 의한 내음성 식물의 특성은 다양한 형태로 나타나므로 식물마다 스트레스 환경과 최적 환경 범위를 도출해 내는 연구는 앞으로도 많은 연구가 필요할 것으로 사료 된다.
엽록소 형광 유도과정 (OJIP) 중 50% 차광 조건에서 무늬사초가 O-J 및 J-I 전이 과정 중 형광량이 가장 낮았다 (data not shown). Marcos et al. (2012)은 수분 스트레스 조건에서 엽록소 형광 유도과정 중 변동형광량 (Vop)이 증가한다고 보고하였는데, 무늬사초와 수호초가 무처리에 비하여 형광량 증가로 인한 O-J 및 I-P 단계에서의 증가 경향이 나타나지 않아 차광에 의한 광이용 효율 저해가 없었다. 따라서 무늬사초와 수호초의 차광에 따른 형태적인 변화는 스트레스에 의한 것이 아닌 생장에 의한 변화임을 알 수 있었다.
지금까지 엽록소 형광매개변수를 이용한 식물의 건전성 평가에 최대형광값/최소형광값 비 (FM/FO ratio)와 광계II 최대 양자수득률을 나타내는 변동형광값/최대형광값 비 (FV/FM Ratio)와 같은 암적응 형광매개변수가 보편적으로 사용되어왔다 (Paillotin 1976; Govindjee 1995, Govindjee 2004; Yoo et al. 2013). 그러나 본 연구의 차광 조건에서 6종의 식물 모두 변동형광값/최대형광값 비 (FV/FM Ratio)와 광합성계 II 전자전달 에너지 플럭스 (ET2o/RC)의 감소 현상은 나타나지 않은 것으로 확인되었다.
그리고 RC/ABS, DIo/RC, RE1o/RC 및 DFtotal ABS 는 차광 조건에서 식물에 따라 증가 또는 감소하는 것으로 나타나 내음성 식물 선발에 활용이 가능할 것으로 판단되며, FV/FM 와 ET2o/RC 는 차광 스트레스 평가를 위한 광화학 생리지표로의 활용이 불가능하다고 판단되었다.
따라서 식물의 형태적 변화와 연계된 광화학학적 특성의 분석 방법은 스트레스 환경의 종류와 범위를 판단할 수 있고, 나아가 형태적인 변화가 생기기 전 스트레스 요인을 제거해 줄 수 있도록 빠른 관리대책을 세우는데 활용할 수 있다.
Conclusion
본 연구는 일조부족, 건조 또는 배수불량과 같은 도시환경에 식재 가능한 지피식물을 선발하기 위한 연구로서 차광 조건 하에서 식물의 생장변화 특성을 살펴보고 일조부족 환경에서 식재 가능한 식물을 조사하였다. 일반적으로 도시환경은 차광 및 건조 등의 스트레스 조건에서 생장하므로 스트레스 내성 식물의 도입이 적절하지만 관상미를 기준으로 식물종을 선택해 왔기 때문에 식재설계에 스트레스 내성 식물이 제대로 반영되지 못하고 있었다. 식물마다 환경변화에 따른 형태적인 변화 특성을 가지고 있는데 이를 판단하기 위한 방법으로 차광 스트레스에 대한 생리적 반응 특성을 분석하였다.
모든 식물에서 차광에 의한 생장의 차이가 나타났는데, 이 중 꽃범의꼬리와 무늬사초, 수호초는 차광환경에서 생장이 더 좋았던 반면, 감국과 노루오줌은 웃자라는 현상이 나타났다. 그리고 엽록소 형광 분석 결과 차광 조건에서 수호초와 꽃범의꼬리, 무늬사초는 광합성계 II 에서 광합성계 I 로의 전자전달 에너지 플럭스가 원활했으며, 광화학적 에너지 구동력이 상대적으로 높게 유지되고 활력이 좋아 내음성 식물로 판단할 수 있었다. 이와 같은 결과는 차광 조건에서 식물의 광이용효율 변화에 따른 광합성 능력의 차이로 해석이 가능하였다.
결론적으로 초장, 마디수, 엽장 등 3종의 생장 특성과 4종의 광화학적 생리지표 (RC/ABS, DIo/RC, RE1o/RC 및 DFtotal ABS)가 도시 환경에 적합한 내음성 지피식물 선발에 활용 가능한 지표인 것으로 판단되었으며, 무늬사초와 수호초가 내음성이 가장 높은 식물인 것으로 확인되었다.
추후 생육조건과 조경식재의 속성정보 연동을 통한 식재설계 수행 시 설계안이 대상지에 미치는 영향력을 객관적 수치로 환산, 분석하고 정해진 기준에 따라 그 결과를 판단할 수 있는 기준으로서 중요하게 활용될 것이라고 사료되었다.
